Open Access
Research Article
Max Screen >>
ISSN: 2576-7623
Copyright: © 2021 Ariff S. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Related article at Pubmed, Google Scholar
Biofilms are complex sessile microbial community extremely resistant to antibiotics. They typically form on both biotic and abiotic surfaces and are usually associated with high rates of mortality and morbidity in patients in hospital settings. Nosocomial infections are the major cause of infections seen worldwide. Bacterial Urinary Tract Infections (UTIs) represent the most common type of nosocomial infection that affect different parts of the urinary tract in both males and females. Staphylococcus species and Proteus mirabilis constitutes 10%-15% UTIs primarily by the quorum-sensing dependent biofilm formation and are highly resistant to antimicrobials. In the current study sulfated polysaccharides were extracted from green algae Chlamydomonas reinhardtii (Cr-SPs), purified by anion exchange chromatography and validated by biochemical and structural analysis. Minimum Inhibitory Concentration (MIC50) of Cr-SPs against S. saprophyticus and P. mirabilis was 760 μg/mL and 850 μg/mL. The time-kill curves and Colony forming unit assays showed bactericidal potential of Cr-SPs in a dose-dependent manner. Cr-SPs inhibited biofilm formation up to 34-100% at 0.5 mg/ml-8 mg/ml in S. saprophyticus and P. mirabilis. Cr-SPs altered the hydrophobic nature of these bacterial cells by ~2-fold as compared to controls inhibiting bacterial adhesion to surfaces. Interestingly, Cr-SPs efficiently distorted preformed-biofilms in a dose-dependent manner. Quantification of total Extra polymeric substance and eDNA of mature biofilms showed Cr-SPs’s ability to destroy biofilms. Scanning electron microscopy analysis of Cr-SPs treated cells showed morphologically altered cells and disintegrated biofilms. Furthermore, Cr-SPs significantly reduced Quorum sensing induced protease and urease enzyme activities and bacterial swimming and swarming motilities. These results indicate the promising potential to develop Cr-SPs as a potential antibiofilm agent against nosocomial infections causing bacteria.
Keywords:Nosocomial Infections; Biofilm Inhibition; Biofilm Eradication
List of abbreviations: Cr-SPs: Chlamydomonas reinhardtii-Sulphated polysaccharides; QS: Quorum sensing; CSH: Cell surface Hydrophobicity
Nosocomial infections (NIs) present a widespread problem in today’s healthcare environment, with between 4% and 10% of hospitalized patients acquiring an infection annually [1]. Nosocomial pneumonias—the leading cause of death in many intensive care units and the second most common NI, account for another 20–25% and are often caused by Gram-negative organism [1]. Various harmful bacteria such as Staphylococcus aureus, Klebsiella pneumoniae, Proteus mirabilis, Staphylococcus saprophyticus, Serratia marcescens, Escherichia coli etc cause most frequent types of hospital-acquired infection such as pneumonia, surgical site infection (SSIs), urinary tract infection (UTIs), and bloodstream infection (BSIs).
Urinary tract infections (UTIs) are one of the most common infectious diseases in humans. The occurrence of these infections is much more frequent in females than in males [2]. Several pathogens including Escherichia coli, Proteus mirabilis, Pseudomonas aeruginosa and Serratia marcescens are often associated in UTIs [3,4]. The uncomplicated UTIs are associated with a variety of organisms, including the Gram-positive bacteria Staphylococcus saprophyticus, Enterococcus faecalis, and other less frequently isolated organisms. If these bacterial infections are left untreated it can lead to various complications such as pyelonephritis, septicaemia, prostatitis, urethritis etc [4]. Therefore, it is essential to treat these infections using various antimicrobial agents. In spite of various available treatments, bacteria tend to develop resistance against antibiotics due to numerous reasons such as target modifications, reduce permeability, target protection, horizontal gene transfer, overuse of antibiotics and biofilm formation [3,5].
Biofilms are one of the major reasons for antibiotic resistance in most of the bacteria. Biofilms are aggregation of bacterial cells adhere to a surface or may remain unattached to any surface and are embedded in a self-producing protective matrix made of Extracellular Polymeric Substance (EPS), which offer strong resistance against antibiotics, host immune response, degrading enzymes etc [6]. According to National Institute of Health (NIH) 60% of all microbial and 80% of all chronic infections are associated with biofilms. Bacteria dwelling inside the biofilm communicate with each other through signalling molecules in response to the population density, the process known as quorum sensing (QS). QS system regulates the expression of several genes responsible for motility, biofilm formation, virulence factor secretion, pigment production etc. Several QS-based factors such as EPS production, swimming and swarming motility have been attributed to the biofilm formation and are associated with UTIs. Generally bacterial cells that grow inside the biofilm are resistant to phagocytes, antibodies and antibiotics, which leads to failure of antimicrobial chemotherapy to clear biofilms [5,6]. An alternative for preventing and eliminating the formation of biofilms of a pathogen is necessary for clinical microbiology.
The sustained failure of conventional drugs in preventing the progress of the disease, coupled with multiple adverse side-effects have driven researchers to look for efficient drug candidates from alternate natural resources [7-9].
Marine ecosystem produces a very rich source of novel compounds with a broad range of pharmaceutical activity. Microalgae are known to produce wide range of diverse secondary metabolites which include alkaloids, polyphenols, terpenes, carbonyls, sulfated polysaccharides, phlorotannins, stilbenes etc that can be developed as alternative antimicrobials agents [10]. Sulfated polysaccharides (SPs) are anionic molecules having a linear structure and are known to have various biomedical applications and also offers varied applications in food, fuel, biotechnological, pharmaceutical and cosmeceutical industries [11-17]. Till now, the antimicrobial properties of marine algae and its compounds were observed but not much work is done related to fresh water algal SPs effect against bacterial biofilms. In the current study P. mirabilis and S. saprophyticus were selected as these are opportunistic bacteria that can cause UTIs, kidney failure, bacteremia, blood and joint infections, skin infections, endocarditis, peritonitis, meningitis and other infections in humans. They are known to contaminate medical devices and implants. These bacteria form firm biofilms and as such there are no known drugs to treat these biofilms [18-20]. Therefore, the aim of the current study is to evaluate the antibacterial, antibiofilm and anti-quorum sensing ability of SPs from freshwater algae against these bacteria. These studies will provide directions to the developments of new generation of natural products which can be used for targeting UTIs causing superbugs.
S .saprophyticus (MTCC 6155) and P .mirabilis (MTCC 425) cultures were obtained from MTCC Chandigarh. Microorganisms were maintained on Growth Medium-3 media (GM-3) with 2 % Agar. For experimental purposes, bacterial cultures were sub-cultured in fresh medium and incubated for 24h. The fresh water algae Chlamydomonas reinhardtii (Cr) was procured from “Chlamydomonas Genetic Centre, Duke University”, USA. The cells were grown and sub cultured on Tris acetate phosphate (TAP) medium. For experimental purposes, colonies are inoculated in liquid media and kept in shaker conditions at a continuous illumination of 300 μmol photons m−2 s−1 as indicated in Sirisha et al. [21].
Extraction and purification of Cr-SPs was carried out as described in Kamble et al. [3]. Briefly, algal cells awre grown to stationary phase. Cells were harvested by centrifuging at 1100 x g for 5 min. The pellet is suspended in 80 % ethanol and macerated with a motor and pestle and incubated in 80 oC water bath for 4 h for extracting SPs. The extract is centrifuged at 4000 x g for 10 min. and supernatant is evaporated using rota evaporator. The elutes were collected and tested for carbohydrate, sulphate and uronic acid contents using standard protocols [3]. The elutes which showed highest carbohydrate content were then pooled, lyophilised and stored at -20 oC. The extract is further structurally characterized by Fourier-Transform Infrared Spectroscopy (FTIR) and Nuclear Magnetic Resonance (NMR) analysis as described in Choudhary et al. [22] and Panigrahi et al. [23].
MIC is determined by subjecting 24 h old cultures to varying concentrations of Cr-SPs ranging from 0.5-8 mg/mL in 96 well plates. After incubation, media was decanted and treated with (3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide) (MTT) to determine the viable cells post Cr-SPs treatment. Cells which are not treated with SPs are considered as controls were maintained. The results were analysed by plotting the concentration of Cr-SPs on X-axis with percent inhibition of bacterial growth on Y-axis [24].
To check if Cr-SPs have bactericidal ability, bacterial cultures were treated with varying concentrations of Cr-SPs for 24 h. After 24 h of treatment, extract was removed, cells were washed with distilled water and fresh medium was added and allowed them to grow for another 24 h. After 24 h of incubation, media was decanted and subjected to MTT dye to determine the viable cells. Appropriate controls were run simultaneously in the plates [24].
For this assay 1010 CFU/mL diluted overnight cultures were used to carry out the time-kill assay. To each well, 100 μL of culture was incubated in GM-3 media along with different concentrations (0.5-8 mg/mL) of Cr-SPs. Plates were incubated at 37 oC with orbital shaking at 120 rpm. Absorbance at 595 nm was recorded every half hour from 0 h till 48 h [25].
The reduction in the colony forming units post Cr-SPs treatment was determined by CFU assay. Overnight culture was diluted to 108 CFU/mL and to each well, 50 μL of culture was added and incubated in GM-3 media supplemented with Cr-SPs in concentration ranging from ½ MIC-8 MIC. Plates were incubated at 37 ◦C with orbital shaking at 120 rpm for 24 h. Post incubation, 50 μL of suspension were spread on GM-3 agar plates and incubated at 37 oC for 24 h and colonies were counted [26].
The inhibitory effect of Cr-SPs on biofilm formation was assessed using the microtiter method. Overnight grown culture was suspended in a fresh medium and Optical Density (O.D) was adjusted to 0.5. To each well of 96-well plates, 100 μL of culture supplemented with GM-3 media was added with different concentrations of Cr-SPs (0.5 mg/mL- 8 mg/mL). After incubation at 37 oC for 24 h, the contents of the well was aspired, rinsed with distilled water (DW), and kept for drying at room temperature (RT) for 10 min. Further, 200 μL of 1 % crystal violet (CV) dye was added and stained for 30 min at RT. The excess stain was decanted, 200 μL of 100 % ethanol was added and O.D were obtained by spectrophotometer at 595 nm [27].
CSH was assessed using Bacterial adherence to hydrocarbons (BATH) assay. Overnight grown culture is taken and re-inoculated in fresh medium and incubated at 37 °C until the O.D. reaches 0.6. The cells are harvested and treated with Minimum Inhibitory Concentration (MIC) and Minimum Inhibitory Concentration (D-MIC) of Cr-SPs and incubated for 24 h at 37 oC at 120 rpm. The cell density is recorded spectrophotometrically at 600 nm. Further to the suspension equal amounts of toluene was added and is vortexed vigorously for 2 min. Then the O.D. of the aqueous phase is recorded. The pre and post recorded absorbance were used for determination of CSH [28].
Overnight grown culture is inoculated in a fresh medium until the O.D reaches to 0.5. To each well, 100 μL of culture along with GM-3 media was added with 15 mM H2O2 and incubated overnight, so as to allow biofilm formation. Next day, to the preformed-biofilms, different concentrations of Cr-SPs (0.5 mg/mL-8 mg/mL) was added along with fresh medium and the plates were incubated at 37 ⁰C for 24 h. After incubation, the cells were stained with 1 % CV followed by the processing of biofilms using 100 % ethanol. O.D were obtained using spectrophotometer at 595 nm [29].
Preformed-biofilm cells were subjected to centrifugation at 5000 rpm for 5 min at 25 oC. Further it was treated with MIC and D-MIC of Cr-SPs for 24 h at 37 oC, 120 rpm and proper controls were maintained. Further the suspension was treated with a mixture of 10 % TCA and equal volume of acetone and incubated at 4 oC overnight. Post-incubation, the mixture was centrifuged at 10000 rpm at 25 oC for 10 min. The weight of treated pellet was compared to the control and these values were used for quantification of EPS [30].
eDNA isolation from preformed-biofilms was extracted by a method reported by Wang et al. [31] with some modifications. In brief, bacterial cultures were treated with 15 mM H2O2 and allowed to form biofilm for 24 h at 37 oC. The preformed-biofilms were treated with different concentrations of Cr-SPs and incubated for 24 h. Subsequently, the plates were chilled at 4 oC for 1 h and treated with 1 μL of 0.5 M EDTA and centrifuged at 5000 rpm for 5 min. The supernatants were discarded and the pellet was suspended in 50 mM Tris–HCl (pH 8). eDNA was extracted with an equal volume of phenol/chloroform/isoamyl alcohol (25:24:1) and with chloroform/isoamyl alcohol (24:1). Further ice-cold 100% ethanol and 1/10th volume of 3 M Sodium Acetate (pH 5.2) was mixed with the aqueous phase in each sample and stored at −20 oC overnight. eDNA was collected by centrifugation at 18,000 X g for 20 min at 4 °C and dissolved in 20 μL of TE buffer. The concentration and purity of the purified DNA were quantified spectrophotometrically by the absorbance ratio A 260/A280.
Approximately 106 CFU/mL of culture was treated with MIC and D-MIC Cr-SPs and incubated at 37 oC for 24 h while maintaining control. 50 μL of these treated cultures was seeded onto 0.3 % Mot medium (10 g/litre tryptone, 5 g/litre NaCl and 0.3 % agar) to assess swimming ability [32]. For analysis of swarming motility 0.5 % GM3 agar plates were used. These plates were incubated for 24 h to 48 h at 37 °C till motility was observed. The images were visualized and recorded using Gel doc [33].
The bacterial cells were treated with MIC and DMIC of Cr-SPs for 24 h. To the supernatant equal amount of 1 % casein substrate was added and incubated at Room Temperature (RT) for 10min. Then 0.4 N TCA was added and incubated at 40 oC for 10 min. The mixture is centrifuged at 5000 rpm for 5 min and 1mL of supernatant is treated with 5 mL of 0.4 M sodium carbonate and 1 mL Folin’s reagent and incubated at 40 oC for 20 min. The protease activity was measured spectrophotometrically at 680 nm [34]..
Cr-SPs treated bacterial cells were centrifuged at 5000 rpm for 5min at 25 oC and the supernatant was used for the assessment of urease activity. Ammonium sulphate was used as a standard. The culture was treated with 2 % urea for 3 h in water bath at 37 oC and then Nessler’s reagent was added followed by incubation at RT for 5min. The absorbance was recorded at 530nm [35].
Glass coverslips were used to grow bacterial biofilms, which are formed after treatment with 15 mM H2O2 from overnight grown culture at 37 °C. This preformed-biofilms were treated with MIC of Cr-SPs and incubated for 24 h at 37 °C with proper controls. After treatment the cells were washed with phosphate buffer saline thrice and fixed for 12 h using 2.5 % glutaraldehyde. Dehydration using graded ethanol (50 %-100 %) was done and the coverslips were subjected to desiccation for 24 h. These biofilms were coated using platinum and visualized using Scanning electron microscope (FEI Quanta 200 (XT Microscope Control) at 20,000 X magnification [36].
The data generated from three sets of each experiments were compared to proper controls and analysed using one-way analysis of variance (ANOVA) using IBM-SPSS software.
The purified extract was composed of 74 % total carbohydrate, ~29 % sulphate, 40 % uronic acid. Further the purified extract is structurally characterized by FTIR analysis which showed side chains characteristic of sulphated polysaccharides [3] and the structural characteristics of Cr-SPs was evaluated by NMR analysis [23]. These results clearly indicated that the extract is indeed enriched with sulphated polysaccharides.
MIC assay showed the dose-dependent growth inhibition of Cr-SPs in these bacteria. The IC50 value was found to be as low as 850 μg/ mL and 760 μg/mL for P. mirabilis and S. saprophyticus respectively. A complete 100 % bacterial inhibition was observed at 8 mg/mL. In case of MBC activity, at 1.6 mg/mL and 1.45 mg/mL of Cr-SPs, there was 50 % bactericidal activity was observed in P. mirabilis and S. saprophyticus respectively. Cr-SPs showed complete bacterial growth inhibition at 4 mg/mL and 8 mg/mL in both the organisms.
The time and dose dependent inhibition of bacterial growth by Cr-SPs was tested using the time-kill assay. The results in Figure 1 shows that Cr-SPs has bacteriostatic effect in both of these organisms. Bacteriostatic effect is seen from 0.5 mg/mL up to 2 mg/mL and a bactericidal effect at 4 mg/mL and 8 mg/mL in a time-dependent manner. When compared to control all Cr-SPs showed a decline in the growth curve directly proportional to increasing concentration of Cr-SPs (Figures 1a and 1b).
CFU results indicate that Cr-SPs inhibited the clonal propagation in both P. mirabilis and S. saprophyticus. It is seen that at concentration beyond 1 mg/mL and 0.5 mg/mL more than 50 % reduction in bacterial colonies were seen in P. mirabilis and S. saprophyticus respectively (Figure 1c and 1d). No colonies were seen at 4 and 8 mg/mL Cr-SPs, depicting the inability of bacterial clonal propagation and survival (Figure e1-e6; f1-f6).
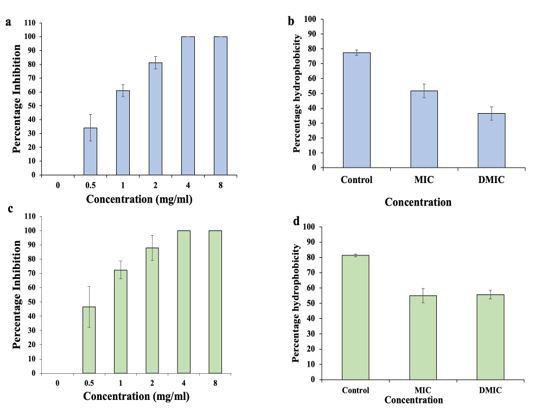
The biofilm inhibition activity of Cr-SPs against P. mirabilis and S. saprophyticus was assessed by CV assay. The results of inhibition assay shows that at ~0.5 mg/mL of Cr-SPs more than ~50 % of biofilm formation was inhibited in both the organisms (Figure 2a and 2c). At 4 and 8 mg/mL of Cr-SPs the biofilms formation in both these organisms were completely inhibited.
Assessment of cell surface hydrophobicity plays an important role in biofilm studies due to its critical role in adherence during biofilm formation. The cells treated with Cr-SPs at MIC and 2MICs showed decreased CSH from ~77 % to ~36 % and ~81 % to 55 % in P. mirabilis and S. saprophyticus respectively (Figure 2b and 2d).
Preformed-biofilms of P. mirabilis and S. saprophyticus were successfully eradicated after Cr-SPs treatment. The results suggested that there was dose dependent eradication of preformed-biofilms. At 0.5 mg/mL of Cr-SPs ~67 % of P. mirabilis biofilm and 55 % S. saprophyticus preformed-biofilms were eradicated. At 4 and 8 mg/mL of Cr-SPs complete distortion of biofilm was observed (Figure 3a and 3d).
The biofilm eradication was further validated by quantification of total EPS content of preformed-biofilms at MIC and D-MIC Cr-SPs. The results depicted distinctive reduction in EPS quantity from control. Cr-SPs (MIC) treated samples showed a 6-fold reduction from control in both these organisms. At 2MIC almost 15-fold and 13-fold reduction in total EPS content was seen in case P. mirabilis and S. saprophyticus respectively (Figure 3b and 3e).
To check if Cr-SPs are targeting and degrading eDNA of the preformed-biofilms, the total eDNA of Cr-SPs treated/untreated samples were quantified. In P. mirabilis the eDNA reduced from 2.6 mg DNA/relative mass in control to 0.03 mg DNA/relative mass at 8 mg/mL. While, in case of S. saprophyticus the eDNA drastically reduced from 1.6 mg at control to 0.02 mg DNA/relative mass at 8 mg/mL (Figure 3c and 3f). A drastic reduction in eDNA with increasing Cr-SPs concentration is in correlation with increasing biofilm eradication and this states the pivotal role of eDNA in biofilms.
Enzymes like proteases and ureases are key players of quorum sensing mechanism that helps in fighting against the host immune systems and establish disease pathogenesis. It was observed that Cr-SPs (MIC and D-MIC) treated P. mirabilis cells showed ~59.51 % and ~56.21 % protease activity in comparison to 100 % in control (Fig. 4a). While, in case of S. saprophyticus cells ~67.63 % and 49.87 % protease activity at MIC and 2 MIC Cr-SPs was observed (Figure 4c).
The urease production in MIC of Cr-SPs treated cells was found to be 63.71 % and 28.91 % and at 2 MIC it was 17.78 % and 8.53 % as compared to 100 % in controls for P. mirabilis and S. saprophyticus respectively (Figure 4b and 4d).
The swimming and swarming motility aids the movement of cells and help in cells to cell communication and disease progression. Inhibiting the motility will help in biofilm disruption at a very early stage. It was observed that P. mirabilis cells treated with MIC and D-MIC of Cr-SPs showed reduced swimming and swarming ability as compared to the controls indicating the potential of Cr-SPs in curbing the bacterial infection (Figure 4e1-e3; Figure 4f1-f3).
The disruption of biofilm as a result of Cr-SPs was visualized by performing SEM analysis. It was observed that the number of adherent cells reduced significantly in the MIC Cr-SPs treated biofilm culture as compared to control. Disruption of biofilm was observed in case of both P. mirabilis and S. saprophyticus. SEM analysis also helped to discover that there is morphological change that occurs in the bacterial cells upon Cr-SPs treatment (Figure 5).
UTIs are among the most common bacterial infection in humans. The financial implications of UTIs are expansive due to its higher incidences [37]. About 80 % of UTIs in humans is caused by various bacteria like E. coli, Enterococcus spp., Proteus mirabilis, Pseudomonas aeruginosa, Klebsiella pneumoniae, Staphylococcus saprophyticus and Enterobacter spp. [37,38]. If these bacterial infections are left untreated it can lead to various complications and death. In spite of various available treatments, bacteria tend to develop resistance against antibiotics due to numerous reasons and one of the main reasons is bacterial biofilms [2,39]. Biofilm is an organized structure caused by aggregation of micro-organisms within a self-produced matrix of extracellular polymeric substance. Biofilms have been noted to cause a damage of over a billion dollars in a year affecting human health and efficiency of industrial products. Hence a lot of research work emphasizes on strategies to combat biofilm production.
The current work focuses on exploring the anti-microbial, anti-biofilm and anti-quorum sensing properties of Cr-SPs. Cr-SPs were extracted and purified by anion exchange chromatography. FTIR and NMR analysis showed the characteristic features of algal SPs [3,23]. Cr-SPs showed efficient anti-bacterial activity against S. saprophyticus and P. mirabilis. The MIC50 of Cr-SPs was found to be as low as 760 μg/mL for S. saprophyticus and 850 μg/mL for P. mirabilis. Cr-SPs also acts as bactericidal agent with 50 % death at 1.6 mg/mL and 1.45 mg/mL in case of P. mirabilis and S. saprophyticus respectively. Time-dependent growth inhibition was observed when the cells were treated with increasing Cr-SPs concentration in range of 0.5-8 mg/mL. Reduction in clonal propagation was observed which was directly proportional to increasing concentration of Cr-SPs (Figure 1). Earlier reports also showed the antimicrobial potential of various marine algal SPs like fucoidan from brown algae, carrageenan from red algae laminarin from brown algae against wide range of bacteria like Pseudomonas aeruginosa, Klebsiella pneumoniae, Vibrio cholera, E. coli, Bacillus species etc. [40-42]. It has been hypothesised that the ability of SPs to inhibit the growth of bacteria is by their ability to permeabilise the bacterial cell membranes forming pores, which causes leakage of intracellular components leading to bacterial death [41].
Cr-SPs showed dose-dependent inhibition of biofilm formation and eradication in S. saprophyticus and P. mirabilis. 0.5 mg/ml of Cr-SPs not only inhibited 50% of biofilm formation (Figure 2) but also effectively eradicated preformed-biofilms in both these organisms (Figure 3) indicating its potential for both infection prevention and treatment. Furthermore reduction in CSH and EPS content clearly aims that Cr-SPs impairs the adherence ability of bacterial cells as well as interacts with EPS to degrade the biofilms (Figure 2b, 2d, Figure 3b and 3e). Earlier reports also showed that SPs from various marine algae showed antibiofilm potential against diverse gram-positive and gram-negative bacteria [43-45]. Antibiofilm screening of fucoidan F85 against dental plaque bacteria showed complete suppression in both biofilm and planktonic mode of growth in Streptococcus mutans and Streptococcus sobrinus [41]. Chmit et al. [46] also showed that polysaccharides extracted from Laurus nobilis effectively inhibited biofilm formation in Staphylococcus epidermis, S. aureus, P. aeruginosa, Enterococcus faecalis etc. Interestingly, in the current study it was observed that with increased concentration of Cr-SPs there was a significant decrease in the eDNA content of the EPS layer in mature biofilms (Figure 3c and 3f). It is known that eDNA plays a crucial role in the development and maintenance of mature bacterial biofilms [47-53]. Earlier studies in S. aureus and P. aeruginosa showed the role of eDNA in matrix formation and stabilisation of biofilms and disruption of biofilms upon DNAse treatment [54].
Quorum sensing (QS) phenomenon is a mechanism that allows pathogenic bacteria to coordinate virulence factors expression for establishing an infection and escaping the host immune response and also helps in in biofilm development in the case of several uropathogens. This signalling mechanism is a system of stimulus to co-ordinate gene expression with other cells and response related to the density of their local population. It depends on the interaction of small diffusible molecules call autoinducers with the receptors of same or different bacteria that initiates transcription of genes responsible for virulence factors [45]. Hence targeting QS molecules can be an effective strategy to prevent biofilm formation and taper the virulence of the organism. Inhibiting QS-induced virulence factors like protease and urease is an important strategy to prevent disease progression [55]. Urease activity is known to cause stone formation and apatites in urinary bladder [56]. Similarly, Protease positive bacteria help to evade host defence mechanism by degrading immunoglobulins thereby initiate infection in the host tissue [57]. Interestingly, in the current study the MIC of Cr-SPs significantly inhibited urease and protease activities in S. saprophyticus and P. mirabilis (Figure 4) respectively. These results were in line with previous reports which showed that various plant secondary metabolites like allicin, curcumin, vanillic acid, phytol, Fluoroquinoles, linoleic acid etc reduced urease and protease activities in Proteus mirabilis, Serratia marcescens, Pseudomonas aeruginosa, Bacillus subtilis etc [58-61]. Our results clearly indicate the potential of Cr-SPs to attenuate quorum sensing pathway thereby helping in controlling the pathogenicity of these UTI bacteria.
In summary, biofilms constitute an important contribution to the high incidence, recurrence, and complications of UTIs. Biofilm research will help in understanding the disease process in a better way and will address in the direction of new and effective prevention and treatment options. Cr-SPs showed promising anti-biofilm properties and can be developed as an alternative therapeutics for combating UTIs (Figure 6).
This work is supported by Department of Atomic Energy, India.
The authors have no conflict in submitting this manuscript.

![]()

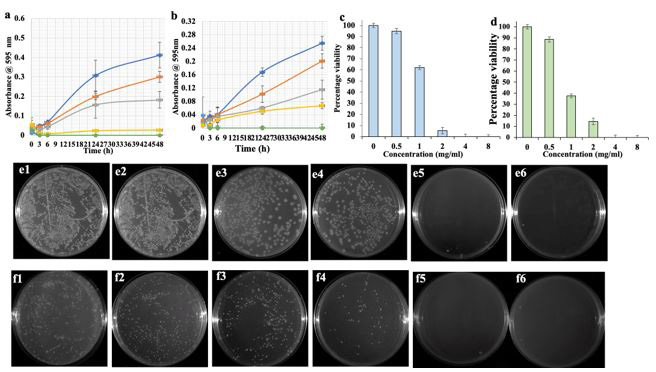 |
Figure 1: Cr-SPs potential to inhibit bacterial growth inhibition over time in (a) P. mirabilis and (b) S. saprophyticus (blue) 0 mg/ml, (orange) 0.5 mg/ml, (grey) 1 mg/ml, (yellow) 2 mg/ml, (green) 4 mg/ml, (light blue) 8 mg/ml. Colony forming unit assay in (c) P. mirabilis and (d) S. saprophyticus after Cr-SPs treatment. (e1-e6)- effect of Cr-SPs on CFU of P. mirabilis. (f1-f6)- effect of Cr-SPs on CFU of S. saprophyticus. Data are the means of three independent experiments ± SE |
Figure 2: Cr-SPs ability to inhibit biofilm formation in (a) P. mirabilis and (c) S. saprophyticus. Cell surface hydrophobicity assay in (b) P. mirabilis and (d) S. saprophyticus. Data are the means of three independent experiments ± SE |
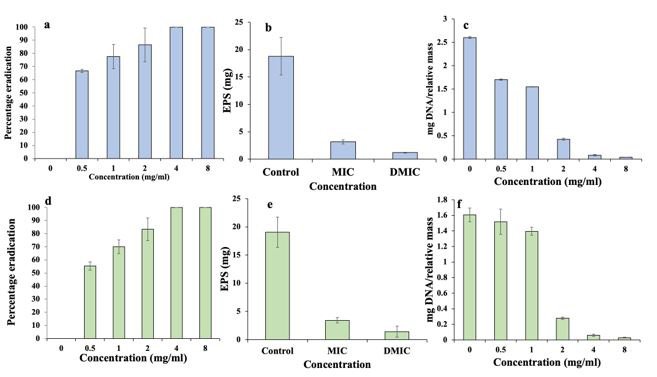 |
Figure 3: Distortion of preformed-biofilms in (a) P. mirabilis and (d) S. saprophyticus after Cr-SPs treatment. Quantification of exo-polymeric substance (EPS) in (b) P. mirabilis and (e) S. saprophyticus after Cr-SP treatment with untreated control. Quantification of extracellular DNA in control and Cr-SP-treated biofilms of (c) P. mirabilis and (f) S. saprophyticus; Data are the means of three independent experiments ± SE |
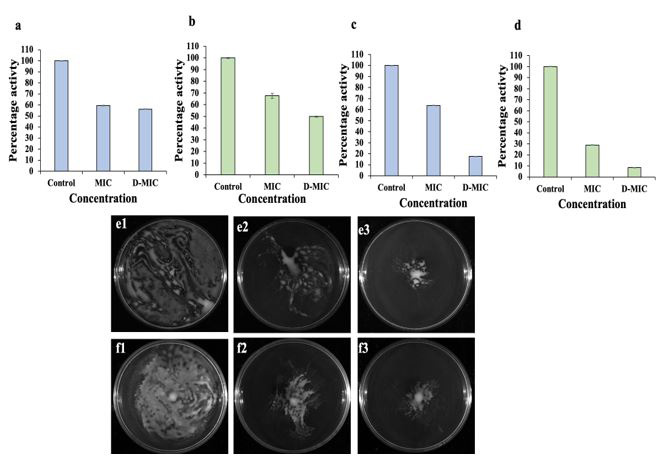 |
Figure 4: Quantification of QS-induced virulence factors. Protease activity assessment in (a) P. mirabilis and (c) S. saprophyticus after Cr-SPs treatment. Urease activity analysis in (b) P. mirabilis and (d) S. saprophyticus post Cr-SPs treatment. Effect of Cr-SPs on (e1-e3) swimming motility and (f1-f3) swarming motility of P. mirabilis. Data are the means of three independent experiments ± SE |
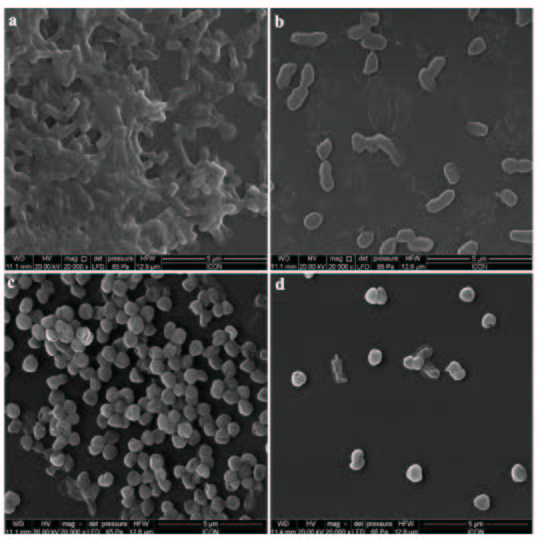 |
Figure 5: Scanning electron micrographs of untreated (a) P. mirabilis and (c) S. saprophyticus, and Cr-SPs (MIC)-treated (b) P. mirabilis and (d) S. saprophyticus biofilms. Magnification × 20,000; bars = 5 μM |
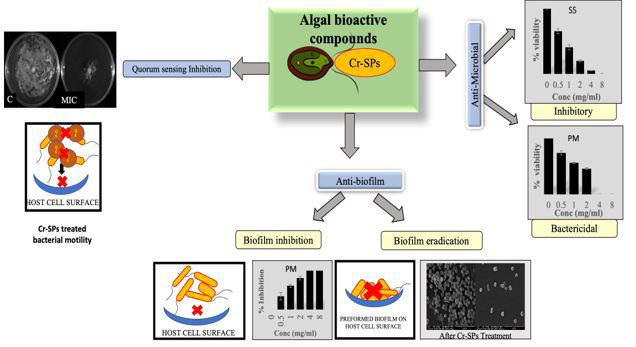 |
Figure 6: Schematic representation of the antibiofilm and anti-quorum sensing potential of Cr-SPs |