Open Access
Research Article
Max Screen >>
ISSN: 2348-9790
Copyright: © 2017 Allingham SM. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Related article at Pubmed, Google Scholar
Rainfall patterns are recognised as important for normal ecosystem functioning in arid environments. Most attempts made to understand the effect of rainfall on amphibian ecology have focused on long-term rather than short-term trends. Here, factors effecting embryonic mortality and clutch size of the African foam nesting frog Chiromantis xerampelina were examined at two ponds from April – June in 2011-2012 in Malawi. A total of 73 foam nests were monitored following spawning. On average 29% of eggs in 2011 and 26% in 2012 suffered mortality from both ponds and this mortality significantly varied between nests. Linear regression showed that the date of spawning had a significant effect on mortality due to moisture requirements of the eggs. The height of nests over the water surface had a negative impact on clutch size and increased rainfall decreased egg mortality in both years at both ponds. In a changing climate, with rainfall projected to become more infrequent in this region, organisms may not be able to rely on rainfall patterns as cues for reproduction, which may have a negative impact on amphibian populations.
Keywords: Chiromantis; Foam nests; Embryonic mortality; Climate patterns
Knowledge on egg and larval survival is essential to understand the population dynamics of amphibians; however, due to the difficulties associated with assessing the survival rate of amphibian embryos and larvae under natural conditions, detailed knowledge of natural survivorship is limited to a few species and populations [1]. Rainfall, oxygen, ultraviolet radiation and temperature can also interact with biotic factors and influence mortality and hatching rate [2-4]. Embryos from amphibian egg clutches are surrounded by a protective perivitelline membrane containing perivitelline fluid and a jelly envelope composed of a variable number of layers [5]. Ambient moisture can influence the physical properties and structure of the embryos and the surrounding protective layers and can potentially alter the ability of embryos to hatch. This is particularly important for the many tropical species known to lay their eggs out of the water, thereby exposing their eggs to variable environmental conditions. Annual and seasonal levels of rainfall are projected to change throughout the twenty-first century [6,7] and as a result, mortality of terrestrial amphibian eggs is predicted to increase in the tropics [8,9]. Most studies considering the effect of rainfall on seasonal growth and survival have focused on seasonal or annual weather patterns [10-12]; but whereas such large-scale rainfall trends can be useful in explaining community level dynamics [13], they may not reveal the specific mechanisms driving individual mortality. In order to elucidate the effect of short term environmental fluctuations on growth and development, rainfall patterns should be considered on a much shorter timescale [2,14].
The African grey tree frog, Chiromantis xerampelina, is a large species of the family Rhacophoridae. Males usually reach 43-75mm and females are around 60-90mm (snout to vent length). This species is mainly distributed from coastal Kenya to the north-eastern South Africa and inland to the North East of Namibia. It occupies a variety of habitats including savannah, shrubland, forests, pastureland and urban areas [15].
The prominent breeding and mating system (polyandry and formation of foam nests) of C. xerampelina appeal to ecologists and has been the focus of attention in many ecological and zoological studies [16-19]. Female C. xerampelina construct foam nests attached to tree branches along the shores of still water bodies where the tadpoles fall and complete their development [15]. The foam of the nest prevents direct observation of the eggs and embryos; therefore, little information is available regarding the embryonic morality and clutch size of C. xerampelina despite the relatively abundant knowledge on many other aspects of the ecology of this species.
Oviposition site selection has been studied in much greater detail in pond breeding species than that of arboreal species and these studies indicate that sites where eggs have been deposited can be influenced by the physical features of a breeding pond [20]. Laboratory and experimental pond observations have shown that both high and low temperatures have detrimental effects on embryos [21-23]. Therefore the timing of spawning may have effects on mortality in the field.
Here, I report about the correlative effect of rainfall on egg mortality of arboreal clutches of C. xerampelina. I also measured clutch size in relation to nest height and date of spawning of this species and discuss a few possible factors causing these variations.
This study was carried out from April 1st to June 5th 2011 and from April 3rd to June 10th 2012 at two ponds in the Vwaza marsh (11º11'15.0 S, 33º27'34.3 E) and the Vinthukutu forest reserve (10º08'46.3 S, 34º11'07.1 E) in Malawi. The Vinthukuzu forest reserve is located close to a lake shore in the Karona district. The reserve consists of dry miombo woodland in transition with partially green understory. The total extent of the reserve is 22km². A pond sized 20 x 15m and 1-50 cm of water depth was chosen as the study target within the forest. The ponds in this forest are characteristically small and shallow and the frogs lay eggs mostly on the emergent vegetation. The Vwasa marsh is seasonal flooded grassland drained by the alluvial plain of the Lewewe River. The Vwasa marsh is consequently deeper than the ponds at Vinthukuzu and the eggs are laid mainly on fringe vegetation and trees near the waterline. The target study area in Vwasa was located in a floodplain near Lake Kazuni. Within the study period, the size of the pond was 25 x 20 m and the water depth varied from 10 to 100 cm.
To identify and quantify the possible factors determining egg mortality, a total of 73 nests were monitored at the Vwasa Marsh and the Vinthukuzu forest. C. xerampelina activity is high throughout this period [15]. The clutches were identified and located the morning after oviposition when the initial number of eggs was recorded. The clutches were subsequently checked twice a day until all the eggs had hatched or died. On the morning of the second day, a series of small containers (10 l) were hung with a small amount of pond water underneath each clutch for monitoring until the emergence of hatchlings [2,24]. When hatchlings began to drop the dead embryos and hatchlings were counted to determine clutch size and mortality.
During each visit, the number of live or dead hatchlings was recorded. Because hatchings fell into small containers below the clutch, missing animals were assumed to be removed by predators. Direct observations of egg predation were also made opportunistically. Submerged clutches, where hatchlings could not be contained by the small container as well as the number of drowned eggs characterized by developmental retardation, was recorded. Halted or impaired development in embryos was indicated by malformation, discolouration or fungal infection.
All analyses were conducted in R version 3.2.3. A linear regression analysis was used to test for a significant relation between egg mortality and the number of days after spawning first spawning. For example the number of eggs that died later on in the season. This analysis was also done for each year separate and then years combined.
A GLM Poisson regression analysis was used to examine the association between the height of the nests and the number of eggs. The analysis was performed for the data from each year separately, and then again for both years combined.
Differences in egg mortality between instances of rain and instances where there was no rain were examined to determine mortality rate. An examination of the distribution of the mortality figures suggested that they had a skewed distribution and were not normally distributed. As a result, the Mann-Whitney test was used to compare between the two sets of figures. This analysis was performed once separately per area and year. The number of eggs collected varied from day to day. As a result, the percentage of eggs that died on each given day was calculated.
A total of 73 nests were found in the study ponds from the 1st April to the 5th June, 2011 and from the 3rd April to the 10th June, 2012. A total of 26,500 eggs were laid in the nests in 2011 and 29,748 eggs were laid in 2012 (56,248 overall). All eggs were attached to tree branches and leaves along the shores of the study areas. The number of eggs per nest ranged from 490 to 1113 (x̄ ± SD = 757.143 ± 215.143; n = 35) in 2011 and from 512 to 1023 (x̄ ± SD = 784.38 ± 173.717; n = 39) in 2012. The average mortality of eggs was 29% in 2011 (x̄ ± SD = 222.30 ± 163.84) and 26% (x̄ ± SD = 197.37.27 ± 154.05) in 2012 from both ponds. In each nest examined, some eggs disappeared after the start of the third day. These eggs may have hatched or been eaten but their fate is unknown. Egg mortality after spawning differed between nests and varied from approximately 20 eggs lost during the first 3 days to 436 after 45 days (Figure 1). The mean mortality for nests selected was 197.94 (SD = 145.40; n = 18) in 2011 and 195.94 (SD = 133.66; n = 18) in 2012. Overall mean for mortality in both years was 196.94 (SD = 137.65; n = 36). Linear regression revealed a significant relationship between mortality and days after first spawning in 2011 (P = <0.01) , 2012 (P = <0.01) and both years combined (P = <0.01).
The height of the eggs over the water surface ranged from 0.50 to 5.6m (x̄ ± SD = 2.89 ± 1.69; n = 73) in both years. By year, the height of the eggs over the water surface ranged from 0.50m to 5.6m in 2011 (x̄ ± SD = 752.51 ± 215.14; n = 35) and from 0.50m to 5.6m in 2012 (x̄ ± SD = 784.38 ± 173.71; n = 39). The height of nests was associated with a significant decline clutch size (Figure 2). This was applied for each year considered separately and also for the data from both years pooled together. There was a significant decline in clutch size in in 2011 (GLM, Z = - 42.05, P < 0.01) 2012 (GLM, Z= - 29.14, P < 0.01) and both years combined (GLM, Z = - 51.15, P < 0.01).
A lack of rainfall after the first 24 hours after oviposition increased egg mortality by 23% in 2011 and 18% in 2012 in Vinthukuzu and by 20% at Vwasa marsh in 2011 and 24% in 2012. The lack of rainfall significantly negatively affected egg mortality in 2011 and 2012 both at Vinthukuzu and Vwasa marsh (Figure 3). Mann Whitney U test indicated a highly significant Increase in egg mortality in eggs that had not been rained on in the first 24 hours in Vinthukutu and Vwasa marsh in 2011 and Vinthukutu and Vwasa marsh in 2012.
The effect that rainfall can have over a species biotic potential in arid ecosystems is well known [25,26]; however, the effect of short term fluctuations on anuran egg survival is less known. This is particularly important for frogs from African savannahs, as rainfall patterns are projected to change throughout the twenty-first century [6].
This study demonstrates the short-term effects of rainfall patterns over tree frog egg survival. C. xerampelina lays its eggs on plants above the water surface and they are consequently highly susceptible to relatively minor variations in environmental conditions. In addition, multiple environmental stressors are more lethal in combination than when considered separately [2]. These results show an increase in mortality following a dry period. In fact, the weather on the days following rainstorms is critical and long rainstorms stimulate spawning.
Large-scale climate fluctuations can have an effect on the ecology of many types of organisms including plants [27,28]. In fact, broad-scale climate patterns have been shown to better predict population dynamics than fine-scale weather measurements [13].
Information on egg mortality in foam nesting frogs is very limited. Mortality rates of 0.183 in R. arboreus [29] lower than that of other foam nesting frogs such as the Malaysian species Polypedates leucomystax (0.340) [30]. Mortality rates of foam nesting frogs are considered moderate compared to other anuran species. For example, the mortality rates reported for several pond breeding species vary from 0.024-0.426 [31-34]. The values reported range between 0.5-0.6 for a tropical hylid species [35]. Failure to fertilize [33,36], desiccation [2] fungal infection [37], low temperatures [38], predation [2] and radiation [39] are all known to increase egg mortality. Fly infestation as been shown to be the main cause of egg mortality in foam nesting frogs [30]. In this study, mortality from desiccation was frequently observed and the height of the nest over the water surface was shown to affect clutch size.
Large rainstorms are projected to become sporadic in this region with climate change [40,41]. Large rainstorms appear to stimulate more frogs to breed, suggesting that they have served as a reliable cue triggering reproduction. Habitat disturbance may interact with the projected less predictable rainfall patterns, potentially affecting egg mortality survival across multiple life stages.
I would like to thank Kayleigh Williams for her valuable advice. I would also like to thank Leila James and Daniel Shepard for their assistance during fieldwork. This research received financial support from Biosearch. Finally I wish to thank the Department of Parks and Wildlife for their support.

![]()

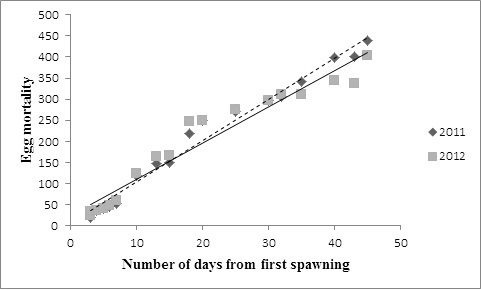 |
Figure 1: Relationship between average egg mortality and days after nest construction during the 2011 and 2012 breeding seasons. Dotted line shows the trend line for 2011 (y = 9.831 + 4.6005; R2 = 0.981, P < 0.01), solid line shows the trend line for 2012 (y = 196 8.9075x + 20.763; R2 = 0.9529, P < 0.01) |
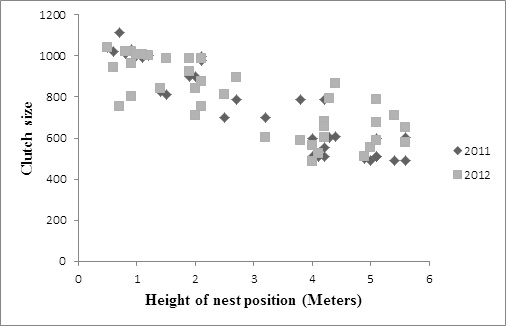 |
Figure 2: An increase in nest height position was associate with a significant decline in clutch size in 2011 (P<0.01) and 2012 (P<0.01) |
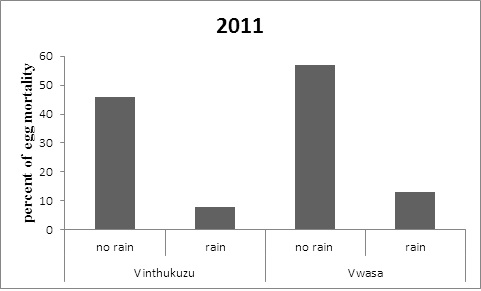 |
Figure 3: Timing of rainfall after oviposition effect on C. xerampelina egg mortality at two ponds. The percentage of total egg mortality increased in clutches not rained on at both ponds in 2012 at Vinthukuzu (P<0.01) and Vwasa Marsh (P < 0.01) and in 2012 at Vinthukuzu (P<0.01) and Vwasa Marsh (P < 0.01) |




































