Open Access
Review Article
Max Screen >>
ISSN: 2575-5498
Copyright: © 2018 Khanh ND. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Related article at Pubmed, Google Scholar
Methicillin-resistant Staphylococcus aureus (MRSA) is one of the greatest fears with the number of serious infections on human health regarding antibiotic resistance. It causes a wide range of infections and bacteremia, ranging from inconsequential superficial skin infections, wound suppuration, even pneumonia or deep-seated tissue infections, which may lead to sepsis and fatalities. The identification of alternative therapeutic compounds has become paramount since there are growing concerns over the extent of antibiotic-resistant pathogenic strains as nowadays antibiotic uses to treat pathogen infections are no longer the golden choices. C. elegans with C. elegan -S. aureus host-pathogen screening platform have been effectively used to screen and identify small molecules as potential anti-virulence agent(s) towards antibiotic-resistant Staphylococcus aureus (S. aureus) as well as to dissect the mechanism of action of the selected molecule(s) as anti-virulence agent(s). The use of this kind of worm with liquid-based assay can be potentially used to simultaneously identify compounds that target host defense as well as bacterial virulence.
Keywords: Caenorhabditis elegans; C. elegans; Liquid-based assay; Anti-infective agents; MRSA infection treatment
Nowadays, the chronic infections leading to serious health problems, caused by anti-biotic resistant bacteria strains, need long-term and complex expansive interventive treatments [1,2]. The clinical infections related to antibiotic resistance have recently been reported increasing not only in the community, but also in live-stock animals [3,4]. The use of antibiotic gradually becomes less useful and less effective now as the morbidity and mortality in human health and animals caused by antibiotic resistance bacteria strains has dramatically increasing with various alarming for several years. The uncontrolled use of antibiotics has led to widespread antibiotic resistance [5]. Each year the number of mortality due to antibiotic resistance in Europe is approximately 25,000 and USA with 22,000 deaths and more than 12,500 deaths in French (Centers for Disease Control and Prevention - CDC) [6]. Human health needs to be improved and protected from infections caused by anti-resistant bacterial strains, so managing and reducing such infections should be carried out as quickly as possible. So far, the priority has been giving to the consideration in finding potential anti-biotic resource to combat the emergence of antibiotic resistance. However, recently it becomes less considerable when the term “anti-infective therapeutic agents” nowadays is turning to new era for human to have neutralized-bacteria healthy life.
There are bacteria strains showing high resistances to antibiotic include vancomycin-resistant enterococci, carbapenem-resistant Acinetobacter baumannii, 3GC resistant K. Pneumoniae, and MRSA. Among those medically-important pathogens, MRSA is one of the greatest fears with the number of serious infections on human health regarding antibiotic resistance. It causes a wide range of infections and bacteremia, ranging from inconsequential superficial skin infections, wound suppuration, even pneumonia or deep-seated tissue infections, which may lead to sepsis and fatalities.
Researchers are on the way to find out an ideal antibiotic in the struggle against resistant-pathogenic bacteria. However, there has no such antibiotic available officially and commercially yet. Available treatments for MRSA infections have been list of antibiotic such as β-Lactam antibiotics, tetracyclines, glycopeptides, macrolides, lipopeptide, aminoglycosides, oxazolidinones, quinolones…[7-9]. The discovery challenges and path forward in finding a novel antibacterial agent which has strongly aptitude to inhibit pathogenic bacteria in almost sites of infection, harmless to the beneficial gut flora and without any undue toxicity to the host. Initially, penicillin was first discovered in 1928, later followed by several highly effective antibiotics. They have been using with usage of drugs as treatment for many bacterial infections. Some of them show wide range of effects in term of dealing with both gram-positive and gram-negative bacteria strains, while some just has effect only one strain [9].
The identification of alternative therapeutic compounds has become paramount since there are growing concerns over the extent of antibiotic-resistant pathogenic strains [10]. This new trend focuses in research for molecules that target the bacterial virulence rather than the bacterial growth. Ideally, new therapeutic paradigms, which can be alternative for conventional antibiotics with great promise, rely on anti-virulence compounds. They have characteristics in selectively disrupting virulence-mediated pathways such as biofilm formation or bacterial quorum sensing. Currently, among different strategies applied for identifying new anti-infectives using whole model organism infection model, C. elegans– S. aureus liquid-based assay has been showing as a highly effective approach [5,11-13]. From that, various conserved innate immune signaling pathways have been disclosed. The use of this kind of worm can be potentially used to simultaneously identify compounds that target host defense as well as bacterial virulence [5,14-16].
From CDC defined criteria, MRSAs are normally classified into community-associated (CA)-MRSA and healthcare-associated (HA)-MRSA and livestock-associated (LS)-MRSA [3,6]. Both CA- and HA-MRSA have geographically wide distributions, therefore worldwide prevalence of MRSA is high [4,7]. They cause several difficult-to-treat infections in human and live-stock animals [3,4]. Consequence by MRSA infections is abundant in both rich and poor countries around the world [17]. Various difficult-to-combat infections caused by this bacterium threaten human life. However, antibiotic uses, which have been mainly used to treat MRSA infections, are no longer the golden choices for those kinds of infections [18]. They act by disrupting the growth cycle of the targeted bacteria strains which causing both nosocomial and community-acquired infections. Result, they might block the synthesis of RNA, protein and other components of bacterial cellular processes. Although these traditional strategies that target cellular viability have proven to be highly effective, they result in substantial stress on the target bacteria, which rapidly selects for resistant sub-populations, leading to resistance towards the antibiotics arsenal [18-20].
MRSA is a developed strain of S. aureus through natural selection and more important under horizontal gene transfer of the mecA gene to at least five distinct S. aureus lineages. It is multi-resistant to β-lactam antibiotics, which include cephalosporins and penicillins (dicloxacillin, nafcillin, methicillin, oxacillin, etc.,). MRSA shows resistance to β-lactam antibiotics because of the expression of mecA gen, which encodes the modified penicillin binding proteins, on staphylococcal cassette chromosomes (SCCmec) [21,22]. There are other expressions of other genes encoding resistance to macrolides or aminoglycosides that can be found on SCCsmec. It consequently leads to limitations in application of antimicrobial therapeutic treatments to those pathogenic bacteria strains [3,22].
S. aureus is well-studied for their ability in acquiring resistance towards antibiotics as epitomized by MRSA and VRSA. The emergence of new and variant S. aureus strains exemplifies the spread of antibiotic resistance. The indiscriminate use of antibiotics, in addition, also reduces the number of beneficial bacteria in the intestinal tract in the host; impairing the host’s immunity therefore the host is susceptible to infections by commensal microorganisms. It is urgent nowadays having a breakthrough for discovery of innovative drugs alternative for antibiotics in combating bacterial infections and impeding the development of antibiotics resistance from bacterial strains.
According to Global antibiotic resistance partnership (GARP) report in 2015, the general situation worldwide on the issue of antibiotic resistance differ country to country and resulted from disease patterns and the use of antibiotics. To be more specific, there is an increase in MRSA infections in India (47%), Latin America, Australia, sub-Saharan (90%). While recently in other countries such as US, Canada, South Africa and European countries, there is a decline in MRSA infections [4,7,23].
The incubation of an individual (both healthy and with compromised immune system) to MRSA infections ranges from a few weeks to many years. When got the infections, initial typical symptoms will progress quickly in the next period of 24 to 48 hours [19]. Living habitats easy to have the emergences of MRSA are hospital, prison, nursing house, school and the bacteria can emerge when people live there contact with contaminated objects such as door handles, floor, dressing, sheet, towel, surface, especially when they have open wounds and weak immune system. As compared to HA-MRSA, CA-MRSA strains show more severe pathogenicity. They not only cause simple surface skin infection but also affect tissues; vital organs even lead to a situation of sepsis, toxic shock and pneumonia [19,22].
LA-MRSA infections are typically transmitted to human from production animals (primarily pigs, then poultry and cattle). This bacteria strain, CC398, is a new variant of MRSA and recently has increased in livestock animals. Interestingly, it is harmless or not shows disease manifestation in food-producing animals but it is severe dangerous to human [3]. A study conducted in 2014 by Mehndiratta and Bhalla showed that CC398 infection occupy more than ¼ in total of MRSA infections in some areas of our planet. Especially, this kind of infection has been found more potentially in group of people working on poultry and cattle industry. MRSA originally has been found in human and then transmitted to animal (livestock and poultry) [24,25]. Interestingly, a contrary result from a single study conducted in Denmark revealed that MRSA infection in livestock and spread to humans [22]. This is credible and is not unfounded because in 2011, a statistic showed that 47% of the meat sold in supermarkets and grocery in US has been contaminated with MRSA. A later analysis was progressed and finally revealed that about 24-52% of these infected S. aureus strains were resistant to some common used antibiotics [26]. There were also other studies conducted in UK and Japan with similar results, MRSA can be found in sold meat products which are pathogenic to consumers [27,28].
Normally, bacterial growth inside the hosts will be inhibited in the presence of penicillin-like antibiotics, which interrupt the synthesis of cell wall materials. However, in case of MRSA infections the bacteria still show their ability to flourish even when treated by such antibiotics [20]. To do so, they have special weapon include mecA and other resistance genes which prevent the activation of the enzyme involved in cell wall synthesis, known as trans-peptidase, despite the destruction from antibiotics [18,20]. Genetically, the resistance genes are involved in extra-chromosomal genetic elements such as plasmids, genomic islands and other genetic elements. The mecA gene is embedded in chromosome, and not coded in plasmid like other β-lactamase genes. Such coding occurs in a genomic island called SCCmec, which shorted for Staphylococcal Chromosome Cassette mec. When MRSA strain acquires this foreign DNA, SCCmec, it can become resistant to methicillin and other antibiotics [20].
There were six types of SSCmec in size of 21 to 67 kb, which containing several resistant genes, have been identified. At least five clones are responsible for the emergence and development of MRSA infections [20]. According to Jensen finding in 2009, different mec and ccr gene they designate into type I-VI of SCCmec. They are type number I, II, III and VIII which normally contain resistant gene can be found in HA-MRSA bacteria [29-31]. In contrast, type IV and V of SCCmec, which do not contain any resistant gene, are normally isolated from CA-MRSA strains [32]. This is resulted from distinction in which SCCmec plasmid they carry. For instance, type I-III of SCCmec carry large plasmids, they cause the bacteria less virulent to their hosts [33]. Antimicrobial resistance rate, type of infections and/or other microbiological characteristics depends much on SCCmec genotype variation [32]. There is another significant difference between HA-MRSA and CA-MRSA strains. HA-MRSA strains normally carry higher cost SCCmec elements, resulting in less virulent expression, more resistant to antibiotics, and they target on hospitalised or immunodeficiency. On contrary, CA-MRSA strains do not carry such high SCCmec elements hence the bacteria themselves express more virulent and toxic to the hosts, targeting on healthy hosts [33].
This mobile genetic element, SCCmec, is not only a genomic island contained mecA resistance gene but also accommodate several genes such as cytolysin gene psm-mec locus, which also confer resistance to HA-MRSA strains against antibiotic uses. The psm-mec belongs to phenol-soluble modulin (PSM) family [31]. It is encoded contiguously to other methicillin resistance genes such as mecA, mecR or mecI. Its peptide, same with other members of the PSM cluster, is capable to lyses human erythrocytes and neutrophils. Hence, these SCCmec elements make the S. aureus resistant to various antibiotics and become one of the most abundantly virulent pathogen [20,29-31].
Initially, the mechanism of the mecA biomarker gene contributing to methicillin and/or other β-lactam antibiotic resistance is known under control of two regulatory genes, mecR1 and mecI genes. The mecR1 gene encodes sensor-inducer while mecI encodes repressor. When the bacteria are being exposed to antibiotics, such as β-lactam ones, the mecI genes in the bacteria itself will be activated and bind to the antibiotics hence forming antibiotic - mecI gene or sensor-inducer. From such binding, it triggers a series of the proteolytic steps that cleaves the mecI repressor, and make it loses its function in binding to mecA promoter [34]. However, the scientists stated that the expression of mecA from mecR1-mecI system occurs slowly and inefficiently; and there are three regulatory genes instead of two previously recognised. Meaning, aside from mecI and mecR1 genes, there has mecR2 recently identified (Figure 1.1). The mecR2 engages with the repressor – called mecI, inhibit its binding to mecA promoter, therefore enables a complete activation of mecA gene for optimal expression of β-lactam resistance, such as PBP2a production. The Figure 1.1 represent for a better and clearer understanding how the three-component (mecI, mecR1 and mecR2) system work [35].
There is another resistant mechanism has been identified with the regulatory genes – blaZ gens in MRSA. Like the mecA gene which under control of two regulatory genes, mecR1 and mecI genes, this blaZ genes also under the regulation by the blaR1 and blaI regulatory genes. The blaI-blaR1 system controls both beta-lactamase and PBP2a production in the bacteria. Both, the mecA and blaZ genes respond for expressions of the bacteria to antibiotics and vary the phenotypic expression of methicillin resistance of the bacterial cells [20]. Since there is a cross-talk between two resistant factors, the regulatory proteins of them, which are mecI-mecR1 and blaI-blaR1, was found to be homologous. Moreover, the mecA system is also initiated by the arrangement of genes coding for blaI and blaR1 and the PBP2a expression can also be regulated by blaI due to the similarity between the operator regions of mecA and blaZ Table 1.1.
Common treatments for MRSA infection are mainly based on tigecycline, linezoid and daptomycin [36]. Nevertheless, this pathogen also showed a concurrently complex increase in resistance mechanism. To be more evident, Snider and Rivard in 2012 reported that the emergence of MRSA strain with high-resistant level and complexity to glycopeptides antibiotic, namely vancomycin and/or teicoplanin, is the consequence of an increase in using of antibiotics. Glycopeptides are antibiotics used to combat MRSA infections since 1960. More specifically, both teicoplanin and vancomycin belongs to glycopepetides antibiotic, but teicoplanin is a structural congener of, and it just has similar activity but a longer half-life, vancomycin. They are less and less used for a variety of reasons [37]. One of the most remarkable reasons is the inconvenient route of administration leading to complicated treatment used for MRSA infections. Since they might show better results when used injections, rather than the normal oral route [38]. Oral consumption of teicoplanin and vancomycin antibiotic, according to clinicians, is not effective and insufficient for controlling systemic infections. Normally, the patients will be treated with vancomycin first, however, when they are vancomycin allergic, the clinicians would suggest using teicoplanin. Furthermore, vancomycin has also showed less effective activity in treatment for MSSA infection than anti-staphylococcal beta-lactam antibiotics [39].
There are other reports also showed the recently treatment failures of vancomycin usage. Meaning, MRSA has been progressively becoming resistant to vancomycin, or vancomycin is no longer a MRSA infection’s treatment choice [40,41]. In 2006, Schito conducted a single study and revealed that, there were growing with several new antibiotic resistant MRSA strains. They have undergone new evolutions and showed high level and complexity of resistance to vancomycin. For serious MSSA infection treatment, Penicillinase-resistant penicillins (dicloxacillin, flucloxacillin) antibiotics with first generation, named cephalosporins, are used to manage the spread or emergences of MSSA [42]. Less serious MSSA infections include skin infection, soft tissue infection; cephalosporins (such as cephalexin, cephalothin, cefazolin) are used as therapeutic treatment. However, this antibiotic cannot be used for all patients because it is allergic to immediate penicillin hypersensitivity such as bronchospasm, angioedema, urticaria [43].
Early, CA-MRSA infections have been treated by using traditional anti-staphylococcal antibiotics such as clindamycin (osteomyelitis), tetracyclines (minocycline, doxycycline) as well as sulfa drugs (sulfamethoxazole/ trimethoprim) [42,44]. Wang and his co-workers in 2006 reported that they successful identified an antibiotic, named platensimycin, used for combatting MRSA. Both HA-MRSA and CA-MRSA showed their resistant level to β-lactam antibiotics [45]. A recent few years, it is believed that vancomycin is the drug of choice for treating of CA-MRSA infections [39,40]. In case of HA-MRSA, it also showed its resistance to these antibiotics and commonly liable to be harmed and extirpated by vancomycin. Later, novel drugs used for MRSA infection treatment were daptomycin and linezolid for a long period of time. In the US, β-lactam antibiotic used to treat MRSA infection symptoms such as skin, soft tissue infections or community pneumonia is fifth-generation cephalosporin antibiotic, kind of β-lactam antibiotic, named ceftaroline [40]. To have the better effect of usage of antibiotics, normally the patients will be assigned to take combined medications of two or even more antimicrobials since the single agent cannot control the pathogenic bacteria [43]. There was also a class of non- β-lactam antibiotic, called oxadiazoles, was successful used to treat MRSA infection in rat models instead [42]. In Australia and New Zealand, the use of newly effective antibiotics, quinupristin/dalfopristin and linezolid (used in antistaphylococcal treatment) is less considered due to its high cost. And it is highly recommended that is only use for a patient who is intolerant of conventional therapy or having infections by high-antibiotic-resistant-bacteria strains [43].
Ceftaroline fosamil is designed to treat MRSA infections and other gram-positive strains of pathogenic bacteria. It belongs to family of cephalosporin antibiotics. This antibiotic also indicated high effect on treatment for community-acquired pneumonia and complicated skin and skin-structure infections [46]. Later in 2010, FDA issued an approval for use of this drug for the treatment of community-acquired bacterial pneumonia as well as acute bacterial skin infections. For skin and skin-structure infections, this drug is effective curing the symptoms as good as vancomycin [47]. Other reports revealed that this antibiotic also can be administered against vancomycin-intermediate S. aureus (VISA) and/or vancomycin-resistant S. aureus (VRSA) successfully.
Clindamycin is a traditional anti-staphylococcal antibiotic with other name osteomyelitis. This drug can be used by oral route or cream to apply on skin. It is indicated that its mechanism is interfering the trans-peptidation process hence inhibiting the protein synthesis process hence killing the bacteria cells [48]. The drug was successful cured some invasive community-acquired MRSA infections [40,48]. It is available in the market with acceptable cost and in form of capsules and oral suspensions. Pain at injection site, rash, diarrhea and nausea are common adverse effects that the treated patients might have. In some case of usage, such as treatment for acnes, it is more successful treated when it is used together with benzoyl peroxide. According to Daum and his study in 2007, it is assigned to cure some cases of MRSA infections [48]. Because this antibiotic is bacteriostatic agent, it could not be applied for all types of MRSA infection [40,49].
Daptomycin is a lipopeptide antibiotic that has been discovered and used since 1980s to treat systemic infections by chronic MRSA and other gram-positive bacteria [40,41,50]. It is recognized with distinct mechanism of action in depolarizing cell membrane and inhibiting protein synthesis in infective bacteria. Meaning, it can cause bacteria cell death by the inhibition of DNA, RNA and protein synthesis [37]. This drug help control MRSA infection, pneumonia which can cause adverse effects for respiratory system in infected patients. According to Figueroa report in 2009, this drug is usually assigned to use with high dosage for sufficient treatments or in other words, the bactericidal activity of this drug is concentration-dependant [51]. Nevertheless, in contrary, another report indicated that using this antibiotic with higher dose may lead to inducing myopathy [40]. More studies and optimization for dose use of this drug is in need to minimize its side effects [37,50].
Linezolid is another antibiotic used to control the spread of both gram-positive and gram-negative strains of bacteria, including MRSA with nosocomical pneumonia symptom [52]. It is assigned to use via oral route by which it could trigger strongest bioavailability and penetration into epithelial lining fluid hence manage or completely cure the pneumonia. Compared to vancomycin, linezolid is better for treatment of MRSA-infected pneumonia. Nevertheless, the common side effects when being used for even short- or long- period could lead to some health problems including nausea, diarrhea, headache, rash or even though symptoms of high blood lactate levels or irreversible optic nerve damage [52,53]. The antibacterial mechanism of this drug based on the inhibition of bacteria protein synthesis which can cause the stop-to-growth or kill the bacteria. Blocking the first stage of protein production of the bacteria make this antibiotic become distinguishable from the others, which are normally interfere at the later steps [53].
Quinupristin-dalfopristin is a combination of two antibiotics of quinupristin and dalfopristin with weighing ratio of 30% and 70%, respectively. This streptogramin drug also has the mechanism of action in killing bacteria cells like other antibiotics such as tetracyclines, clindamycin and tigecycline by interrupting the bacterial protein synthesis. It is designed for MSSA infections [49,54]. Although the side effect with severe myalgias is unexpected, in some cases when MRSA infections cannot be controlled by other antibiotics, it was assigned to use this combination antibiotic [54].
Another combination antibiotic is trimethoprim-sulfamethoxazole which combines two antibiotics, trimethoprim and sulfamethoxazole with ratio of one-five respectively. This drug is available in both for-used ways including oral and intravenous [55]. The mechanism of action of this drug is suppressing folic acid synthesis of the bacteria leading to additive effect in inhibition of bacterial cell growth [40,56]. There are considerable side effects from using this drug such as nausea, colic, diarrhea, nasan, mental depression, dizziness, uremia-developing patients [56]. According to the report by Liu and his co-workers, this drug is not being approved by FDA for MRSA infection treatments; it is being used to treat non-invasive community-acquired MRSA infections. A comparison in anti-bacterial effect have been made by a clinical study with the result showed that Trimethoprim-sulfamethoxazole antibiotic is less effective than others such as vancomycin and tetracyclines [40].
Telavancin is a bactericidal lipoglycopeptide antibiotic used to treat gram-positive infections, including MRSA. It is a semi-synthetic derivate of vancomycin drug. Hence, the mechanism of bactericidal activity for therapeutic treatment also based on the inhibition of cell wall synthesis and/or depolarisation of bacterial membrane. Since 2009, FDA approved this drug for skin-structure and complicated-skin infection. Currently it is used to treat MRSA and vancomycin-intermediate S. aureus infections [57-60]. Accidentally, it was found with the ability to cure the infective endocarditis (a hardly-to-treat disease with biofilm formation) though it was not initially assigned for this kind of infection’s patient. Therefore, it is suggested that it has part in biofilm-related infections [58,59]. Remarkably, it is recommended not to use for pregnant women because it can cause teratogenicity and this was report since 2011 by FDA for the adverse effects [57].
Tetracyclines are also bacteriostatic agents which can cause the inhibition of protein synthesis of the pathogenic bacteria by blocking the attachment of charged aminoacyl-tRNA to the A site on the ribosome [61]. They belong to a big family of antibiotics; include minocycline and doxycycline [40]. In case of the MRSA strain carries tetK gene, exhibiting drug out the cells, tetracyclines antibiotic might become less effective [62]. Among members of tetracyclines, minocycline has the most effective mechanism of action as compared to others. the reason explain for it could be that, it restricts to a minimum of resistance mechanism of MRSA strains due to its longer half-life, better absorption and enhanced tissue-penetration [40,61]. It was also found with higher effects than other antibiotics including tigecycline, linezolid, daptomycin and even vancomycin in controlling MRSA isolated from biofilm [63]. There are many side effects of minocycline such as fatty liver, hepatitis, tinnitus, breathing complications, growth of bone in fetus. Remarkably, due to the adverse effects of minocycline in MRSA treatments, it is not highly recommended or assigned for patients with drug-induced lupus-like-syndrome, renal disorders, autoimmune and pancreatitis or in pregnancy stage [61,63].
Tigecycline is an antibiotic used to treat both gram-positive and gram-negative pathogenic bacteria. This is indicated to treat skin-structure or complicated-skin, community-acquired pneumonia and intra-abdominal infections [37]. Like the mechanism of bactericidal activity of daptomycin and/or telavancin drugs, it also causes bacterial cell death by prohibiting bacterial protein synthesis. The reports showed that, this drug was designed with specific structure with a modified side chain to minimize major bacterial resistance [37,49]. Clinical studies were conducted and recommended that tigecycline is effective treatment for serious MRSA infections. This drug is used via intravenous infusion and no available options for oral route intake. Noticed, there were precautions with use of this drug due to increase in mortality risk of patients (cause is unknown clearly) in 2010 by FDA [57]. It must be careful when using this antibiotic for pregnant ladies and children. FDA then also issue a regulation about precaution part in the drug label due to this increase in death risk as compared to other alternative treatment. The warning was made after a clinical trial in total of 7400 patients with serious infections caused by MRSA because the statistics indicated higher risk of all-cause mortality with those who use tigecycline instead of other appropriate treatments [57,64].
Even though there were many reports revealed that ratio of infections caused by MRSA have scaled down in recent years, the needs to search out potentially applicable alternative therapeutic treatments for those infections remain high [65]. The reason explains for it could be that, MRSA still exist as one of the most common single multi-drug resistance bacterium. Recently, there are several studies about therapeutic treatments used for MRSA infections. Most of pathogenic S. aureus strains show the antibiotic resistant level to penicillin, and commonly spread from hospital into the community [43]. In 2010, Sherlock and colleagues tested and indicated that, honey contains some specific components can be used to kill MRSA [66,67]. Another in vitro study conducted later revealed that, the extracted compounds from cannabis plant, common hemp, called cannabinoids and pinene can be used to combat MRSA infections [68]. Extracts from oak also showed anti-bacterial activity in combat to MRSA [69]. Most of these in vitro need more downstream works to publish anti-bacterial molecule(s) use for therapeutic treatment against MRSA infections.
From all the above mentioned and discussed, MRSA has developed its own resistance mechanism to many in-used antibiotics such as tetracyclines and vancomycin. Evidentially, the more in usage of antimicrobial agent, the more complex resistance mechanism the pathogenic bacteria strains acquire. Moreover, precautions with considerable number of adverse effects as part of toxicity remain in these antibiotics. FDA issued approval list of drugs for MRSA infection treatment (Table 1.2) [57]. From the table, we have clearer view of available antibiotics for therapeutic treatments caused by MRSA such as complicated intra-abdomen infections, bacteremia infective endocarditis, nosocomial pneumonia. Therefore, the current mission of scientists in finding new potential anti-infectives replaceable for older drugs is necessary to completely treat and/or control MRSA infections.
C. elegans belongs to Animalia kingdom, Nematoda phylum, Chromadorea class and Rhaditida family. The use of non-vertebrate tractable host models (such as Drosophila melanogaster, C. elegans, Dictyostelium) to study virulence mechanisms of human pathogenic bacteria strains as well as the defense mechanisms from the host has become more popular recently. C. elegans, an in vivo model genetic host, has been used, complementary to those already in existence to establish a novel host-pathogenic model system to discover potential therapeutic anti-infective molecules [5,10,16,70]. In other word, using C. elegans as a host model for MRSA pathogenesis, researchers decipher the mechanisms of host-pathogen interaction through the understanding of MRSA or MSSA infection. Previously, the traditional platforms for drug discovery were normally in vitro screening and the potential compounds then were tested in a whole and completed organism they showed poor pharmacokinetic activities and toxic. Interestingly, using C. elegans for anti-infectives discovery to initially test the compounds has many advantages and over those previous limitations [16].
There are serious limitation with conventional mammalian models (rodent, primate, …) in determining the in vivo effects of the test compounds such as ethical constrains, time-consuming and laborious procedures, high cost. In addition, mammalian models have normally been used for finding potentially new drugs leads, nevertheless they are strictly prohibited to use for screening large number of compounds. Those make the progress of finding new antimicrobial agents becomes more slowly. With the use of C. elegans, these limitations can be overcome as it can be used to conduct the initial testing of compounds in a whole animal system. Screening of compounds with the nematode whole animal model also allows preliminary assessment of drug toxicity as worms treated with these compounds will show similar or decreased survival when compared to the untreated control [71]. The ability of self-fertilization of the worm allows us to use it in automated high-throughput drug screening method with low cost and rapid cultivation of large population without any ethical concern [16]. Moreover, the worm is not only help detect potential antimicrobial compounds that directly target the pathogen viability, but also with the compound that alter pathogen virulence or enhance the host immune response. However, this model organism cannot completely replace mammalian models because of its favorable temperature of growth and development, its efficient detoxification system which limits the capacity to identify compounds that act via modulation of host defenses [72]. Hence, there is a need to investigate more with this promising host model organism used in drug discovery.
This worm is small (~1mm length), has short generation time, simple anatomy and fully sequenced genome. Working with the nematode has no concerns or worries either for genetic and genomic knowledge or laboratory techniques, all are available and very easy-to-manipulate. Because C. elegans is hermaphrodite, it can self-fertilization and allow for a very-easy-to-get big population hence it allows for preliminarily screening of toxic or non-toxic compounds without any ethical constraints. The transparent body allows them to be visualized under the microscopes both in normal or to-be-fluorescent conditions [16]. In nature, this worm/metazoan uses soil bacteria as food source [70]. This reveals that they have protective responses combat pathogens themselves. Hence it is up to the widely use of these microorganisms as an attractive host–pathogen model [73]. The worm itself has physical protective tools against microorganism intrusion such as cuticle and grinder. Notwithstanding, these physical defense is not protectively perfect because a few bacteria can penetrate deep into the worm’s intestines through the grinder’s worm then cause infection and consequently kill the worms. Then they can proliferate inside the gut and produce toxins that can kill the worm [73-75].
There were several publications conducted using C. elegans, the soil-borne transparent nematode, as non-mammalian host to study the host-pathogen interactions [5,70,73,76-80]. Mallo et al. in 2002 conducted microarray experiments with infected C. elegans and from that identified some genes that may play a role in innate immune response. Their report was the first profiled defense-related genes for the worm responses to bacterial Serratia marcescens infection by using high-density cDNA microarray. Detail, there were remarkable upregulation of the expression of any genes which also can be found in the immune system of other model organisms and encode lysozymes and lectins [81]. First profiling of defense-related genes to antimicrobial agents, later many studies have been conducted to identify the conserved innate immunity signaling pathways against the pathogens which are quite similar among the nematode defense system and other mammalian. Apart from identifying the virulent factors which cause the worm death [82], some other reports also successfully revealed pathways associated with the defenses system of the worm including transforming Growth Factor-b (TGF-b) pathway by Mallo et al, 2002 [81], the Insulin/Growth Factor-1 (IIS) pathway by Garsin and his co-workers in 2003 [83] and later the p38 Mitogen Activated Protein Kinase (p38 MAPK) pathway by Troemel et al. in 2006 [84].
C. elegans has been used in various in vivo screening platforms as a host model to study and identify the resistance mechanism of the worms with Pseudomonas aeruginosa (P. aeruginosa)and S. aureus infection [85]; Lactobacillus acidophilus (L. acidophilus) strain NCFM in mechanism enhancing gram-positive immune responses [86], Escherichia coli and S. aureus causing infections in diabetes [87], S. aureus infection [88], identify new bacteriocin to combat infections from S. aureus strain RF122 causing severe disease such as mastitis [89]. Those reports have contributed more insights about pathogenic-host model interactions among C. elegans and pathogenic bacteria strains.
From those basic of knowledge, the researchers have conducted studies using C. elegans for anti-infective discovery (Table 1.2). It is another remarkable advantage of using C. elegans in vivo host model. In 2014, Kong and her colleagues were successfully used C. elegans platform for anti-infective discovery. From this report, they found two compounds extracted from Orthosiphon stamineus (O. stamineus) leaf and Nypa fruticans (N. fruticans) root in total of 29 synthetic compounds and 37 natural extracts that showed anti-infective properties. They showed ability in prolonging the C. elegans from S. aureus infection without affecting bacterial cell viability which means. The hits from this report may also be further extended to higher organism or even in human [10]. However, the worm is not a perfect host model and cannot completely replace other higher animals in screening anti-microbial candidates due to some limitations need to be acknowledged such as suitable environmental temperature for both maintenance and screening period (must not be exceeded 37 °C and in range of either 16, 20 or 25 °C) [16]. Moreover, potential compounds that act via modulating host defenses are not easy to be identified because this worm has an efficient detoxification system itself [72]. The utility of this nematode to screen anti-infectives molecules for therapeutic treatment against S. aureus infection should be pursued and need more intensive studies [10,72].
Liquid-based assay using C. elegans as a Host Model-Effective Screening Approach for Anti-Infective Compounds
Conventionally, the antimicrobial compounds/molecules which can be used for bacterial infections were probed using in vitro cell-culture based assay. It is less effective as compared to the later one called in vivo pathogen- host platforms of screening. The reasons for it could be that, there was a lack of pathogen-host interaction which serves as reliable proof for further test in higher organism and even in human. Later, in 2006 there was an alternative approach for anti-infective high-throughput screening which was conducted by Moy and his colleague. The experimental procedure is set out by using 96-wells plate with C. elegans-Enterococcus faecalis platform for screening [11]. This is a liquid-based screening assay hence the worm has a constant time to be exposed in the liquid media instead of agar surface. The compounds both natural extracts from marine and plant or synthetic ones were tested whether they can rescue the worm from E. faecalis infection. The results showed that using this infection model, the researchers can screen a large number of promising compounds for antimicrobial with very low minimum inhibitory concentration (MIC) lower than 31 mg/mL. They showed capability in curing the worm survival and some can inhibit the replication of the bacteria [71]. From that initial research, there were many others work on and looked forward to finding novel anti-infectives and they have achieved some remarkable hits 16.
An infection model C. elegans -S. aureus screening platform has been conducted by Kong and her co-workers in 2014 to probe novel compounds/small molecules can be effectively used to combat MRSA infections [10]. Finding from Sifri and his team in 2003, there is a difference between infection caused by E. faecalis and S. aureus to the worm, the second is just transient and not persit in the worm gut [82]. The liquid-based screening assay was applied with total of more than 70 compounds from natural extracts, marine and from the synthetic ones. Seven best candidates showing potential anti-microbial effect screened from them then were subjected to MIC determination assay. Finally, they found the best effective anti-microbial molecule is the curcumin extracted from Curcuma longa [10,82].
Another experiment in 2014 by Rajamuthiah et al. also used C. elegan -S. aureus host-pathogen screening platform to screen anti-infectives that can be used to combat MRSA infection. The liquid-based assay was used to probe for 640 drugs for MRSA infections which approved by FDA for other therapeutic treatments. From that, closantel, inherently used for intestinal worm treatment in human and veterinary, was probed for prolonging the survival of MRSA-infected C. elegans. Meaning, this drug can be potentially applied for MRSA infections with very low MIC. It also showed capability in dealing with other gram-positive and gram-negative strains of bacteria with low concentrations. For VRSA infection, this drug showed effective control only with concentration of 0.78mg.mL-1. Moreover, this screening platform is more strongly believed effectively used for probing anti-infectives candidates because from that study, the drugs which have been used as antibiotic combating MRSA so far also probed as hits from the results of screening [90].
The advantages of using liquid-based assay rather than the conventional agar-based assay include cost effective, simple, time efficient, high sensitivity, and with low concentration of MIC.
Anti-Infective Compounds-Potentially Novel Alternative Therapeutic Treatment for MRSA Infections
Anti-infective can be defined as an agent that is able to prevent or counteract infections based on their capability to inhibit the dissemination of an infectious agent or directly kill the pathogen. Depends on the mode of action the antimicrobial agents, for instance the antibiotics, perform their bacteriostatic or bactericidal properties, by which they inhibit bacterial propagation or kill the bacteria, they can be categorized accordingly (Kong et al. 2016). Because they target on bacterial viability, hence their actions lead to the substantial stresses on the targeted bacteria strain and their anti-biotic resistance. To overcome these pitfalls, researchers are trying to find out other therapeutic treatments for bacterial infections. These alternative approaches must satisfy the request for not affect the viability of the bacteria and may strengthen the host innate immune responses and host survival (Figure 1.3).
Nowadays, with the widely used of C. elegans as a host model in C. elegans - S. aureus infection model, the anti-infectives with novel potential characteristics have been identified with very low of MIC which allow for further test in higher animal and human. The advantages of using C. elegans, a very useful host model for drugs discovery and development nowadays, in in vivo screening force for the brighter future with no more subsidiary on antibiotics to combat MRSA infections.
The use of the whole animal C. elegans model helps investigate anti-virulence therapeutics that target virulence factor(s) expressed in vivo when host-pathogen relationship is intact. Moreover, identification of novel compound(s) as anti-virulence could strongly support for further development as anti-pathogen therapeutics. Also, the understanding of bacterial virulence mechanism in vivo and potentially novel virulence factors as anti-virulence targets. Using liquid-based assay rather than the conventional agar-based assay has more advantages including cost effective, simple, time efficient, high sensitivity, and with low concentration of MIC. From my review on the use of C. elegans model as well as liquid based assay, I thereby would like to recommend this approach as it will help researchers screen and identify small molecules as potential anti-virulence agent(s) towards antibiotic-resistant S. aureus as well as to dissect the mechanism of action of the selected molecule(s) as anti-virulence agent(s). The use of this kind of worm can be potentially used to simultaneously identify compounds that target host defense as well as bacterial virulence.
The author would like to thank Vietnam International Education Development–Ministry of Vietnam Education and Training and Dong Nai Technology University–DNTU for funding this paper review by scholarship under the Project 911 (2016-2020).

![]()

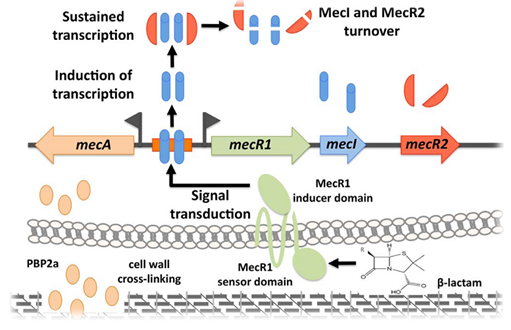 |
Figure 1.1: Three-component system of the mecA regulatory locus. In the presence of a β-lactam antibiotic, mecR1 is activated and rapidly induces the expression of mecA and mecR1-mecI-mecR2. The anti-repressor activity of mecR2 is essential to sustain the mecA induction since it promotes the inactivation of mecI by proteolytic cleavage. In the absence of β-lactams, mecR1 is not activated and a steady state is established with stable mecI-dimers bound to the mecA promoter and residual copies of mecR1 at the cell membrane [35]. |
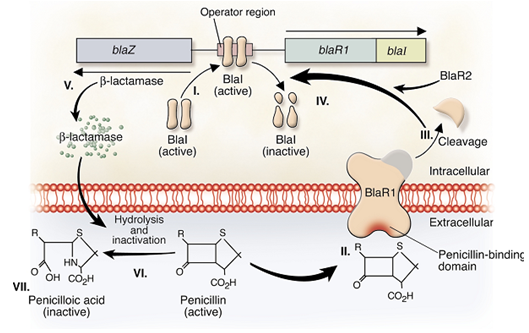 |
Figure 1.2: Induction of staphylococcal β-lactamase synthesis in the presence of the β-lactam antibiotic penicillin |
 |
Figure 1.3: Possible results from C. elegans anti-infective screen |
Antibiotics |
Resistance gene(s) |
Gene product(s) |
Mechanism(s) of resistance |
Β-lactams |
blaZ |
Β-lactamase |
1.Enzymatic hydrolysis of Β-lactam nucleus |
Glycopepetides |
Unknown |
Alterned peptidoglican |
1.Trapping of vancomicin in the cell wall |
Quinolones |
parC |
1.Parc (or grlA) component of topoisomerase IV |
1,2.Mutations in the QRDR region reducing affinity of the enzyme-DNA complex for quinolones |
Aminoglycosides (e.g., gentamicin) |
Aminoglycosides-modifying enzymes (e.g., aac, aph) |
Acetytransferase, phosphotransferase |
Acetylating and/or phosphorylating enzyme modifying aminoglycosides |
Trimethoprim-sulfamethoxazole (TMP-SMZ) |
Sulfonamide: sulA |
Dihydropteroate synthase |
1.Overproduction of p-aminobenzoic acid by the enzyme |
Quinupristin-dalfopristin (Q-D) |
Q: ermA, ermB, ermC |
Ribosomal methylases |
1.Reduce binding to the 23S ribosomal sununit |
Table 1.1: Mechanism of S. aureus resistance to antibiotics used [20] |
|||
Drugs/ infections |
Complicated SSSI |
CAP |
Nosocomial Pneumonia |
Bacteremia |
Infective endocarditis |
Complicated intra-Abdominal Infections |
Vancomycin |
✓ |
✓ |
✓ |
✓ |
✓ |
|
Linezolid |
✓ |
✓ |
✓ |
|||
Daptomycin |
✓ |
✓ |
✓ |
|||
Tigecycline |
✓ |
✓ |
✓ |
|||
Telavancin |
✓ |
|||||
Ceftaroline |
✓ |
✓ |
||||
SSSI: skin and skin-structure infections; CAP: community-acquired pneumonia There are absences of doxycycline, quinupristin-dalfopristin, minocycline, clindamycin and trimethoprimsulfamethoxazole because they have not been approved by FDA with indications for use to treat MRSA infections yet. |
||||||
Drugs/ |
Description |
Infection Model |
References |
Berberine derivatives |
Berberine is a natural alkaloid found in plants. |
C. elegans-E. faecalis |
Tomkiewicz et al. 2010 |
Celecoxib |
An FDA-approved non-steroidal anti-inflammatory drug that blocks the COX-2 enzyme. It is used to relieve fever, pain and inflammation. |
C. elegans-MRSA |
Thangamani et al. 2015 |
Artilysins |
A modified form of endolysins, enzymes produced by bacteriophages to hydrolyze the bacterial cell wall, with improved outer membrane-penetrating capability |
C. elegans-P. aeruginosa |
Briers et al. 2014 |
Curcumin |
Main active component Extracted from Curcuma longa |
C. elegans-S.aureus |
Kong et al. 2014 [10] |
Lamotrigine |
An FDA-approved anticonvulsant drug that also inhibits ribosome biogenesis in bacteria, thereby preventing the growth of bacteria. |
C. elegans-Salmonella enterica |
Stokes et al. 2015 |
Auranofin |
A form of gold complex that is mainly implicated in the treatment of rheumatoid arthritis |
C. elegans-S. aureus |
Fuchs et al. 2016 |
Table 1.3: In vivo screening using C. elegans as a host model: probe anti-infective molecules/compounds for anti-microbial effect [16] |
|||




































